
Молекуларлық биология биологиялық, физикалық және химиялық әдістер жиынтығын кеңінен қолданады, олардың кейбіреуі оған алдыңғы ғылымдардан (биохимия, цитология, генетика және т.б.) «мұра ретінде» қалдырылса, ал басқасы оның Молекуларлық объектілермен жұмыс жасау үрдісінде өздігінен дамуының арқасында пайда болды. Төменде ең көп қолданылатын тәсілдер тізімі берілген және олардың қолданылу салалары келтірілген.
Микроскопия – оның ерте тарихы бар, ол XVII ғасырдан басталады, 1611 жылы Й. Кеплер жарық сәулесі микроскобын жасау үрдісін ұсынады, ал А. Левенгук соның көмегімен алғаш рет ,бір жасушалы бактерияларды зерттейді (1638). 0,4-0,7 мкм аралығын қамтитын осы сәуле микроскопиясы М. Шлейден мен Т. Шванның 1838 жылы жасушалық теорияны жасауына мүмкіндік берді, бұл теорияға байланысты ядросы бар жасуша – барлық жануарлар мен өсімдіктер құрылысының құрылымдық және функционалдық бірлігі болып табылады. Сосын, 1857 жылы Р.А. Колликер алғаш рет митохондрияға, ал В. Флемминг – митоз кезіндегі хромосомдардың мінездемесіне түсініктеме берді.
Интерференциондық (1930), фазалық-қарама-қарсылық (1932) және ақыр соңында, электрондық (1939) микроскоптарды жасау жетістіктері микроскопия дамуының ең маңызды деңгейі болып табылады
Әдетте, өлшемдері 20 Å (2 нм) объектілерді ажырата алатын және қазіргі замандағы үлгілерінде шегі 0,1 нм болатын электрондық микроскоптар вирустар, жасушааралық органеллдерді, белоктық-нуклеиндік комплекстер (хромотин, рибосомдар, информосомдар) құрылымын және кейбір белоктық молекулаларды зерттеуде кең қолданылыс тапты. Осы әдістің нұсқаларының бірі – криоэлектрондық микроскопия – қазіргі кезде рибосомдардың нәзік құрылымын зерттеуде басты рөл атқарады.
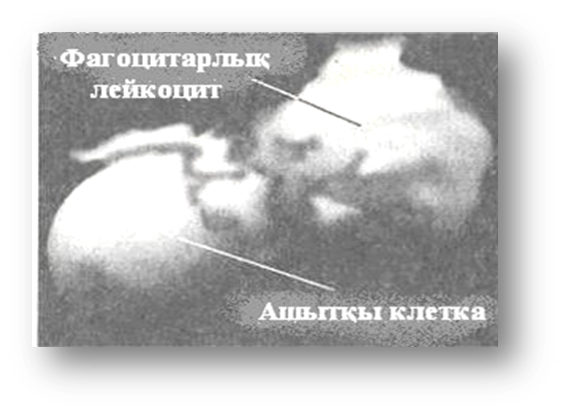
Сканирлеуші электрондық микроскоптар жасуша құрылысының ең үлкен әсерлі және ақпараттық үшөлшемдік (көлемдік) суреттерін алуға мүмкіндік береді (1-сурет).
1-сурет. Электрондық микроскопия әдісімен алынған суреттердің мысалдары:
А – фаг бөлшектерінің (×200 000) электрондық микрофотографиясы. Солдан оңға қарай: икосаэдр түріндегі фаг – φХ174, табак мозаикасының вирусы (таяқша бейнесіндегі бөлшек және фаг Т 4 ; Б – сканирлейтін электрондық микроскоп көмегімен алынған фагоцитоз көмегімен ашытқылық клетканы жұтушы полиморфты-ядролық лейкоцитттің фотографиясы
Рентгендік-құрылымдық талдау – рентген сәулелерінің дифракциясы негізінде құрылған (толқын ұзындықтары 10 -10 м электромагниттік сәуле шашырату); ол молекулалардағы атомдардың үшөлшемдік орналасуын анықтауға мүмкіндік береді (0,1 нм-нен төмен аралықта). Бұл әдісті Англияда Г. Брэгг пен Л. Брэгг жасап шығарды және соның көмегімен ақуыз, ДНҚ және РНҚ молекулаларының құрамы туралы іргелі мәліметтер алынды.
Қазіргі кезде бұл әдіс алынған мәліметтерді компьютерлік талдаумен қоса, биополимерлердің үшөлшемдік құрылымын зерттеуде басты әдіс болып табылады.
Радиоактивтік изотоптар – тірі жасушалардағы нуклеин қышқылдары, ақуыздар, углеводтар және басқа да молекулаларды зерттеуде кеңінен қолданылады. Радиоизотоптар тұрақты емес және зарядталған бөлшектер – электрондар бөліп шығаратын таралу әсерінде болады немесе гамма-шашырау пайда болады. Ал жартылай ыдырау периоды өте қысқа 14 тәуліктен ( 32 Р изотобында) өте ұзақ ( 14 С изотобында) 5570 жылға дейін созылады. Электрондарды сцинцилляциялық есептегіш не Гейгер есептегішіндегі газдардың иондануы бойынша, немесе радиавтография әдісімен (олардың сезімтал фотоэмульсия қабатындағы күміске әсері бойынша) анықтайды.
Радиоактивтік молекулалар әртүрлі жасуша аралық процесстерді зерттеу кезінде: молекулаларды синтездеген кезде, молекулалардың жасуша ішіндегі ықшам орналасуын, олардың жасушадағы және бөлек компартменттердегі жұмыс жасау уақытын, макромолекула-лардың кейбір бөлшектеріндегі химиялық айналуын және т.б. анықтаған кезде қолданылады. Егер, мысалы, жасушаны РНҚ-ның радиоактивтік ізашарымен (3Н-уридинмен) инкубацияласақ, РНҚ-ның алдымен ядрода синтезделетінін, сосын жасушаның цитоплазмасына өтетіндігін анықтауға болады. Радиоактивтік белгіні зерттелетін молекулаларға ферменттер арқылы енгізуге болады. Осылайша фосфотрансфераза мен нуклеотидилтрансферазаны қолдана отырып, ақуыздар, көмірсулар және нуклеин қышқылдарына, АТР әртүрлі радиоактивтік формалары арқылы белгі енгізуге болады (2-сурет).
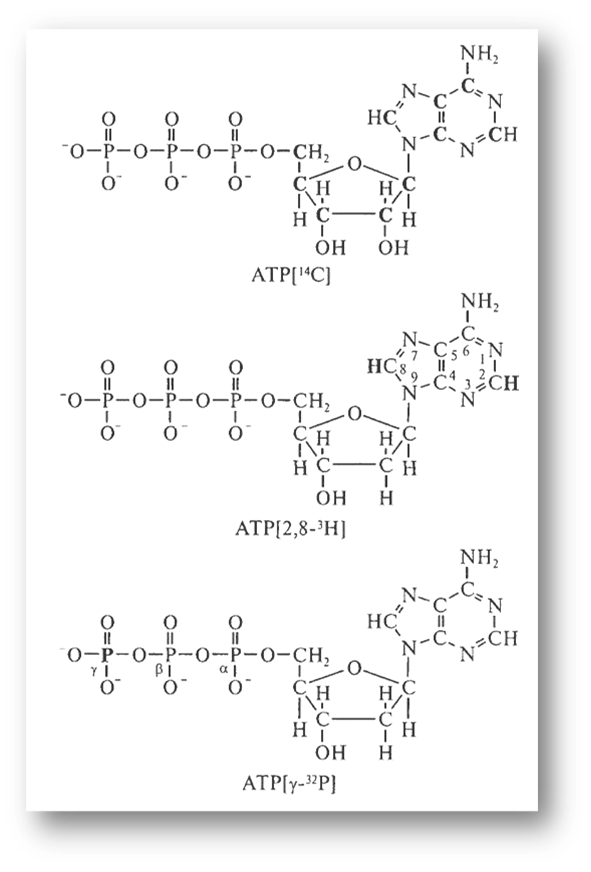
2-сурет. АТР радиоактивтігінің үш түрі.
Радиоактивтік атомдар қара шрифтпен көрсетілген. Радиоактивтік атомдардың орналасуы мен түрлерін көрсететін белгілері келтірілген
Ультрацентрифугалау (седиментациялық талдау) – 1926 жылы Т. Сведбергтің аналитикалық ультрацентрифуганы жасап шығарып, соның көмегімен алғаш рет гемоглобиннің Молекуларлық массасын (68 кДа) анықтауынан кейін өте кең таралды. Бұл әдісте седимента-ция (тұнбалау) жылдамдығы бөлінетін компоненттердің өлшемі мен түріне байланысты анықталады және S – седиментация коэффициен-тімен өрнектеледі. Седиментация коэффициенті секундпен өлшенеді және мына формуламен анықталады: (dx/dt)/w 2 x, мұндағы, х – айналым центріне дейінгі қашықтық (айналу осінен центрифугалық пробирканың түбіне дейінгі қашықтықпен анықталатын центрифуга роторының радиусы, см -мен өлшенеді); dx/dt – седиментация жыл-дамдығы (см/с); w – ротор айналымының бұрыштық жылдамдығы (рад/с). Седиментация коэффициенті әдетте, өте аз мәнде болады, сондықтан Сведберг бірлігімен өлшенеді (S): 1S = 1*10 -13 c. Гемогло-бин молекуласының седиментация коэффициенті – 4,5S, тРНҚ молекуласынікі – 70S, лизосоманікі – 9400S.
ХХ ғасырдың 40-50-жылдары А. Клод пен Ж. Барше жасуша орга-неллдерін бөлу үшін дифференциалдық центрифугациялау әдісін ой-лап тапты, соның көмегімен де Дюв (de Duve) 1953 жылы алғаш рет лизосомаларды, содан кейін пероксисомаларды бөліп шығарды. 1957 жылы М. Месельсон, У. Сталь және Дж. Виноград нуклеин қышқы-лын хлорлы цезий тығыздығының градиентінде центрифуга тәсілімен бөлу әдісін ойлап тапты, соның көмегімен ДНҚ репликациясы жартылай консервативті жолмен жасалатыны белгілі болды (7-тарауды қараңыз). Ультрацентрифугалау әдістерінің әртүрлі нұсқалары жасушаішілік компоненттер мен макромолекулаларды (нуклеин қыш-қылдары мен белоктар) бөліп шығару үшін, сондай-ақ олардың молекулярлық массалары мен седиментация коэффициентін анықтау үшін кеңінен қолданылады.
Әртүрлі молекулаларды, ең алдымен, ақуызды, нәзік фракциялау мен тазарту үшін қазіргі кезде алуан түрлі физика-химиялық тәсілдердің көптеген топтары қолданылады. Ақуызды фракциялаудың кеңінен таралған: хроматография, электрофорез және изоэлектрлік фокустеу әдістерін қарастыралық.
Хроматография – орыс ғалымы М.С. Цвет алғаш рет ашқан әдіс, ол 1906 жылы өсімдік жапырақтарының боялған сығындыларын бор үгіндісі бар тізбекте фракциялады.
Қазіргі кезде хроматографияның алуан түрлері бар, онда ақуыздарды зарядтары бойынша (ион айырбастау хроматографиясы), молекулалардың өлшемі бойынша (жиі гель-фильтрация деп аталатын гель-хроматография) және матрикспен алдын ала біріктірілген заттардың белгілі бір химиялық группаларымен ерекше өзара әрекет жасау қабілеті бойынша (аффиндік хроматография) ажырататын әртүрлі типтегі матрикстер (тасымалдаушылар) қолданылады. Хроматографияның сан алуан түрлерінің ішіндегі ең тиімдісі аффиндік хроматография (тектілік хроматографиясы) болып табылады, оның негізінде өзара әрекеттесетін заттардың молекулаларының өзара ерекше қарым-қатынасы (бір-бірін тану) жатады. Мысалы, фермент субстраттарын алдын ала матрикспен біріктіріп, ферментті тізбекте ұстап тұруға, сосын, қалған белоктар («балластық») бөлініп шыққан соң, оған гомогендік жағдайында элюирация жасауға болады. Матриксты ерекше антителалармен («қарама-қарсы денелер») жабдықтап, бұл тасушыларды осы антителалар танып білетін белоктарды бөліп шығаруға пайдалануға болады. Ал матрикспен ДНҚ фрагменттерін біріктіріп, хромосомалардың белгілі бір бөліктерімен ерекше байланысатын белоктарды бөліп шығаруға болады.
Сонымен қатар, жоғары тиімділікті сұйықтық хроматография (HPLC) да кең таралды. Бұл әдісте диаметрі 3-10 мкм микросфера түрінде гомогендік ортаны құра алатын және жоғары қысымда діңгек арқылы ерітіндінің бірқалыпты ағуын жүзеге асыратын, әдейі жасалынған кремний-органикалық смола қолданылады, осының нәтижесінде, хроматография процессі аздаған минуттарда жүргізіледі және талдау жасалынатын молекулалар қоспасының сезімтал фракциялануын қамтамасыз етіледі.
Электрофорез – бұл әдістің негізінде, белгілі бір оң немесе теріс зарядтардың қосындысына ие белоктардың электр өрісінде зарядтар өлшеміне және молекулалардың формасына сәйкес орын ауыстыру қабілеті жатады. Электрофорезді су (буферлік) ерітіндісінде жүргізуге болады, бірақ, әдетте, оны әлдеқандай полимерлік тасушыларда: крахмалдық, агароздық немесе полиакриламидтік гельдерде, целлю-лозалық немесе нитроцеллюлозалық пластиналарда және т.с.с. жүргізеді.
Электрофорездің ең қарапайым целлюлоза пластинкаларындағы тәсілі 1956 жылы В. Ингрэмнің белокты трипсинмен өңдеу нәтижесінде алынған адам гемоглобинінін, протеолиз ферменттерімен ыдыратылған фрагменттерін фракциялаған кезде қолданылды.
Осы әдістің көмегімен пептидтік карталар – «фингерпринттер» (бармақ дақтары) алынды, нәтижесінде орақ түріндегі жасушалық анемияның β-тізбегіндегі гемоглобинде бір ғана амин қышқылы басқа амин қышқылымен ауыстырылғанда пайда болатыны анықталды.
Қазіргі кезде ақуыздарды бөлу үшін полиакриламидтік гелде (ПААГ) жүргізілетін электрофорез әдісі жиі қолданылады, ол көл-денең тігістерінің саны жоғары және бақыланатын инерттік матрикс түрінде жүргізіледі. Гел ұңғыларының өлшемдерін өзгерте отырып, Молекуларлық массалары бойынша бір-бірінен әлдеқайда өзгеше (бірнеше он мыңнан жүздеген мыңға дейінгі дальтон) белоктарды фракциялауға болады. Бұл әдісті 1959 жылы С. Раймонд ойлап тапты, ал соңынан Б.Дэвис пен Л. Орнстейн оны әрі қарай жетілдірді. Ол молекулярлық массалары мен зарядтары әртүрлі ақуыздар қоспасын нәзік фракциялауға мүмкіндік берді. 1966 жылы Дж. Майзель ұсынған ПААГ-тағы натрий додецильсульфатымен электрофорездеу осы әдістің одан әрі модификациялануы (жетілдірілуі) болып табылды.
Натрий додецильсульфаты (ДСН, SDS, ДС-Na) күшті иондық детергент болып табылады, ол ақуыз молекулаларын бір созылған тізбекте ашып, оларға артық теріс зарядтар береді (3, А – сурет). Осындай (денатурацияланатын) гелде өзінің нативтік формасын жоғалтқан ақуыздар, олардың полипептидтік тізбегінің өлшемдерімен (массасымен) ғана детерминацияланатын жылдамдықпен оң зарядталған электродқа (анодқа) қарай жылжиды. Бұл жағдайда гель Молекуларлық елек рөлін атқарып, ұсақ полипептидтерді оңай өткізіп, ал одан ірі молекулалардың жылжуын баяулатады, ал ол, өз кезегінде ақуыздарды олардың Молекуларлық массаларына сәйкес бөледі (3, Б – сурет).
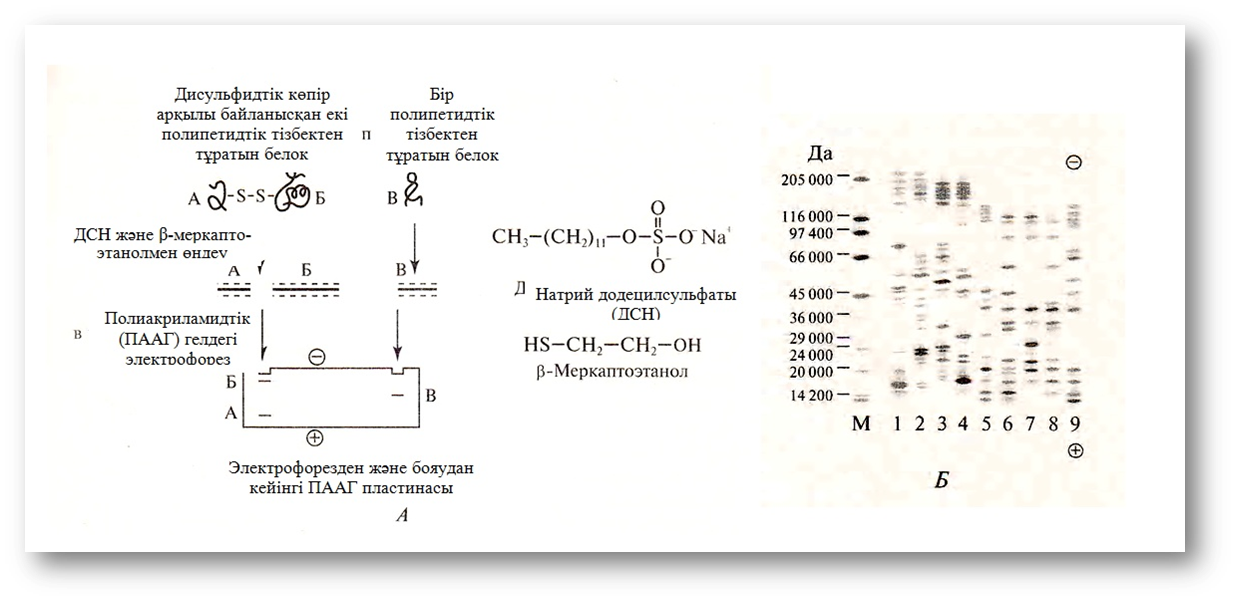
3-сурет. Белоктарды натрий додецил-сульфатымен электрофорездеу:
А – бөлек полипептидтік тізбектердің арасындағы байланыстарды (дисуль-фидтік) үзетін, белок глобулдарының қопарылысына әкелетін және оларға артық теріс заряд беретін белоктарды ДСН және β-меркаптоэталонмен өңдеу (электрофорез кезінде бұндай денату-рацияланған белоктар полипептидтік тізбектерінің өлшеміне (массасына) кері пропорционал жылдамдықпен анодқа қарай орын ауыстырады); Б – полиокриламид гелінде ДСН-мен электрофорездеу тәсілімен картоптың әр түрінің жапырақтарының ерігіш белоктарын фракциялау нәтижесінде алынғанэлектрофореграмма: 1-9 – картоптың түрлері; М- Молекуларлық массаның маркері.
SDS-пен электрофорездегенде, әдетте, ақуыздар суббірліктерінің арасындағы дисульфидтік байланыстарын үзетін (немесе қалпына келтіретін) β-меркаптоэталонмен өңдеу қолданылады, ол мультимер-ақуыздардағы суббірліктердің саны мен массасын анықтауға мүмкіндік береді. Электрофорезден кейін ақуыздарды Кумасси бояуларымен немесе күмістендірумен (минорлық ақуыздар) айқындайды, ол электрофореграммаларда белоктың өте аз мөлшерін (10 нг) анықтауға мүмкіндік береді.
ХХ ғасырдың 70-жылдарында Швецияда изоэлектрофокустеу деп аталған ақуыздарды электр өрісінде бөлудің жаңа әдісі ашылды. Электрофоретикалық буферлік ерітіндісімен белоктар рН-тың белгілі бір мәнінде фракцияланатын электрофорездеу әдісінен оның айырмашылығы – изоэлектрофокустеу кезінде ақуыздар арнайы реагенттер (амфолиндер) арқылы жасалатын рН градиентінде бөлінеді. Изоэлектрофокустеу процессіндеақуыздар электр өрісінде тек молекулалар зарядына қатаң сәйкестікпен жылжиды және рН градиентінің олардың изоэлектрлік нүктесіне сәйкес келетін нүктесінде (рІ), яғни молекулалары электрлік бейтараптанатын нүктелерде тоқтайды (фокустенеді) (4-сурет).
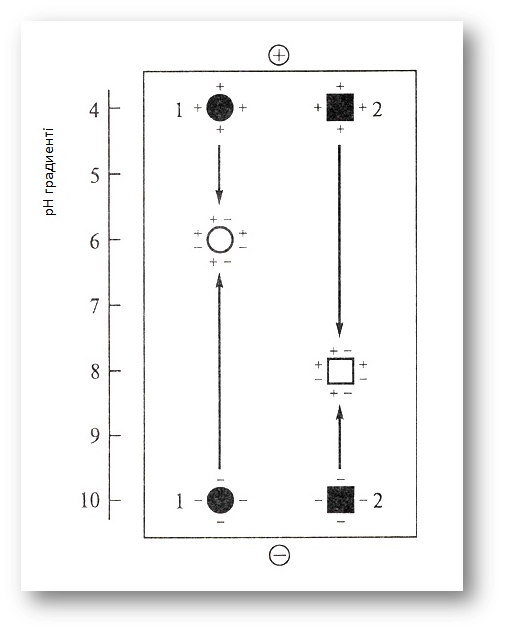
4-сурет. Изоэлектрофокустеу әдісі бойынша белоктарды бөлу.
рН төмен болғанда белоктар оң (+) оқталады, жоғары рН - теріс (-). Электр өрісінде белоктардың молекулалары қарама-қарсы оқталған полюстерге қоныс аударады да, изоэлектрлік нүктелерге (pI) сәйкес рН градиенті нүктелерде тоқтайды. 1-ші және 2-ші белоктардың изоэлектрлік нүктелері тиісінше рН 6,0 және 8,0 болады.
Изоэлектрофокустеу белоктарды зарядтары бойынша бөліп қана қоймай (рІ) бойынша рН-тан 0,005 бірлікке ғана айрықша болатын ақуыздар бөлінеді), олардың изоэлектрлік нүктелерін тез және нақты анықтауға мүмкіндік береді.
1975 жылы П. Офаррелл ойлап тапқан ПААГ-та SDS-пен электрофорездеу мен изоэлектрофокустеудің үйлестірілген әдісі – екіөлшемдік электрофорез атағын алды. Бұл процедурада ақуыздар-ды толық еріту, зарядтарын өзгертпей, полипептидтік тізбектерді денатурациялау және диссоциациялау үшін алдымен β-меркапто-этанолмен және мочевинаман өңдейді. Содан соң, ПААГ-та изоэлек-трофокустеумен ақуыздарды зарядтары бойынша бөледі, сосын оларды ПААГ-та SDS-пен электрофорездеу блогына (перпендикуляр бағытта) әкеледі, онда ақуыздар Молекуларлық массасына қарай бөлінеді.
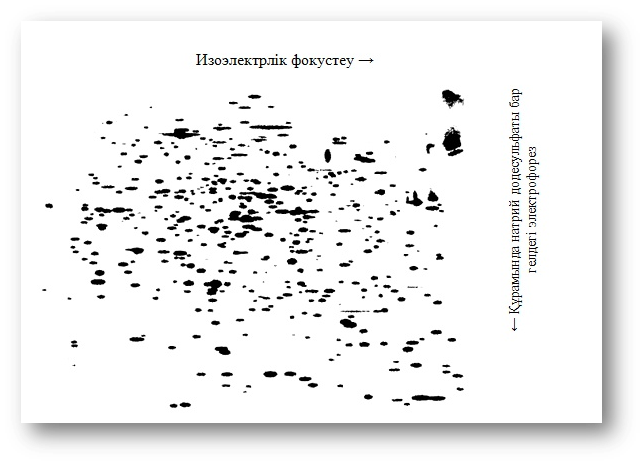
5-сурет. Ішек таяқшаларының белоктарын 14 С-аминқышқылы қосылған екіөлшемдік электрофорез әдісімен бөлунәтижесін бейнелейтін радиоавтограмма Пластинаға фотографиялық пленканың полиакриламидтік гелін басу арқылы алынған радиоавтограммада белоктардың полипептидтік тізбегінің әр бөлігіне сәйкес 1000-нан астам дақты байқауға болады.
Сонымен, алдымен зарядтары, ал содан соң өлшемі (массасы) бойынша бөлуді байланыстыра отырып, бір мезгілде 2000-ға дейін полипептидтік тізбектерді бөлуге, яғни бактерияның барлық белоктарының көпшілігіне талдау жасауға болады.
Жасушалар өсіндісі – бұл әдіс тауық эмбрионының өзінің өмірге бейімділігін тұзды ерітіндіде жақсы сақтайтындығы алғаш рет анықталған, 1885 жылдан басталады. 1907 жылдан бастап жіпшелер (ткань) фрагменттерінің дақылдары (эксплантанттары) қолданыла бастады. Қазіргі уақытта бір типті клеткалар алу үшін жасуша культурасын диссоциациялау қолданылады. Кейде клетка культурасында , шексіз көбейе алатын және жасушалық сызықты құрайтын мутанттар пайда болады.
Үзіліссіз бөліне алатын рак жасушаларынан олардың айырмашылығы, мутанттық жасушалар, кейбір затардың бетімен жанасқан кезде, жақсы өседі. Жасушалық сызықтар көптеген бір типті клеткалардың негізі болып табылады және 70°С-да өзінің пролиферацияға қабілетін жоғалтпай ұзақ уақыт сақтала алады. Жасушалалық сызықтар ең көп қолданылатындары тышқандар мен атжалмандардың фибробластарының, сонымен қатар, эпителиальдық адам жасушыларының сызықтары. 1952 жылы HeLa сызығы ретінде кеңінен мәлім, қазіргі кезге дейін қайтадан өріліп келген адам жатырының карцинома жасушалық сызықтары алынды.
Клеткалық линиялар біртектілігін клондау арқылы жоғарылатуға болады. Клон – бұл бір клетка-ізашардан таралатын клеткалар популяциясы. Клондау әдісінің көмегімен, мутациясы белгілі бір гендерге әсер ететін мутанттық клеткалық линияларын бөліп шығарады. Бұндай клеткалардың белгілі белоктары бойынша кемістіктері болады, ал ол қалыпты клеткалардағы оның функциясына талдау жасауға мүмкіндік береді.
Екі жасушаны бір-бірімен қосу арқылы гетерокариондарды – екі бөлек ядролы жасушаны алуға болады. Митотикалық бөлуден кейін гетерокарион, барлық хромосомалары бір ядрода бірігетін, гибридтік клеткаға айналады. Осындай гибридтік клеткалар әртүрлі жас бөлек хромосомаларының функцияларын, клетка ішіндегі компоненттерін (митохондриялар, ядролар және т.б.) зерттеуге мүмкіндік береді. Оларды клондап гибридтік клеткалық линияны алуға болады. Гибридтік клеткалар тұрақты болмайтыны анық. Жеке алғанда, адам-тышқан гибридтік клеткалар адам хромосомасын бірте бірте жоғалта бастайды, ал ол осылай олардың генетикалық картасын жасай отырып, кейбір хромосомалардың функциялары туралы жорамалдауға мүмкіндік береді.
Клеткасыз жүйелер – белгілі бір Молекуларлық процесстерді зерттеу үшін таптырмайтын жүйе, себебі клеткаларда өтетін әртүрлі қажетсіз реакцияларды болдырмайды. 1954 жылы П. Заменчик белок биосинтезін (трансляция) зерттеу үшін алғашқы клеткасыз жүйені алды. Осы жүйелердің мүмкіндіктері, әрі қарай, А.С. Спирин, М. Номура тағы басқа зерттеушілер трансляция бөлшектерін зерттеген кезде тамаша қолданыс тапты. Мұнда клетка экстрактарынан белок синтездейтін жүйелердің маңызды компоненттері (мРНҚ, рибосомдар, тРНҚ және т.б.) бөлініп шығарылды, сосын оларды бірте-бірте жүйеге енгізіп, әрбір компоненттің белок синтезіндегі рөлін анықтады. Клеткасыз жүйелердің көмегімен генетикалық код айқындалды, ол үшін мРНҚ ретінде белгілі құрамдағы синтетикалық олиго- және полинуклеотидтер қолданылды. Клеткасыз жүйелер, сонымен қатар басқа да маңызды Молекуларлық-генетикалық процесстерді: ДНҚ репликациясы мен транскрипциясын, РНГ сплайсингісін және т.б. процесстерді зерттеген кезде пайдаланылады.
Моноклоналдық антителалар (бөтен денелер) – күрделі қоспалардағы молекулаларды анықтаудағы (идентификация) өте сезімтал құрал, сондықтан олар Молекуларлық биологияда кеңінен қолданылады. Антителалар негізінде, бөтентекті қоспалар – антигендерден қорғану үшін омыртқалылардың бөліп шығаратын белоктары (иммуноглобулиндер) болып табылады. Омыртқалылар-дың клеткалары антиген бөлшектерімен бір-бірінен ерекше антителалардың өте көп форма түрлерін (10 8 ) синтездей алады. Атап айтқанда, осы антителалардың ерекшелігі, клеткалардағы молекулаларды айқындаудағы олардың бірегей мүмкіндіктерін анықтайды. 1976 жылы В-лимфоциттерді клондау әдісі жасалынды, ол антителалардың белгілі бір түрін ғана секрет түрінде бөледі, соның арқасында осы антителаларды көптеп алуға болады. Бірақ дақылдағы В-лимфоциттердің тіршілігі қысқа, сондықтан оларды «мәңгі» ісіктегі В-лимфоциттерден шығатын клеткалармен қосады. Пайда болған қоспадан дақылдарда дамитын және белгілі бір антителаларды синтездейтін гибридтер алынады. Бұл гибридтік клеткалар гибридомдар деп аталады. Оларды бөлек-бөлек клондап әрбіреуі моноклональдық антитела көзі болып табылатын клондар алынады. Моноклональдық антителалар бір ғана клеткадан алынып, белоктар үшін абсолюттік ерекшеліктері болады: олар аминқышқыл қалдықтарының 5-6 тобының конфигурациясын айқындауға қабілетті. Антителалардың әр түрін, әрі қарай белоктардың клеткалардағы орналасу нүктесін анықтау үшін және белоктарды аффиндік хроматографияда тазарту үшін зонд ретінде пайдалануға болады. Жақын болашақта моноклональдық антителалар каталитикалық белсенді молекулалардың жаңа топтары – абзимдерді жасаудың көзі болып табылуы мүмкін (2-тарауды қараңыз).
Каталитикалық белсенді белоктар (ферменттер) – биохимиялық зерттеулер әдістерінің ең маңызды құралдары. Олар Молекуларлық биологияда кең қолданыс тапты және оларды пайдалану мысалдары келесі тарауларда келтіріледі.
Бақылау сұрақтары:
I-тарау
Молекулалақ биология әдістері
1.Молекулалық биология қандай ғылымдардың жетістіктерін өз зерттеулерінде қолданылады?
2.Микраскопиялық зерттеу тәсілдері қай уақыттан бастап қолданылып келеді?
3.Микраскоптың қандай түрлері малекулалық биологияда қолданылып жүр?
4.Рентген құрылымдық талдау неге негізделген?
5.Радиоактивті изотоптар қандай мақсаттағы зерттеулерде қолданылады?
6.Жартылай ыдырау периоды деген не, және әртүрлі биоэлементтердің ыдырау периодын көрсет?
7.Аналитикалық ультрацентрифугалау әдісі неге негізделген?
8.Молекулалық биологияда хроматографияның қанндай түрлері қолданылып жүр?
9.Электофорез әдісі неге негізделген?
10.Клеткалық жүйелер қандай мақсатта қолданылады?